Methods
MICRO-CARD-FISH
CAtalyzed Reporter Deposition Fluorescence In Situ Hybridization combined with MICROautoradiography is currently the method of choice to address the activity of selected prokaryotic groups. Fluorescence in situ hybridzation with cyanine 3 labeled probes already has a long history, however, this approach frequently suffered from low detection rates especially in open ocean settings with low ribosomal content of the prokayrotes.
Probes for CARD-FISH are labeled with a horseradish peroxidase (HRP) tag. After hybridization of the probe to the target site a signal amplification step is introduced where tyramides labeled with FITC or CY3 conjugates are deposited near the hybridized probe through the enzyme activity of the horseradish. This makes it possible to detect prokaryotes with very low ribosomal content and detection efficiencies of up to 80%.
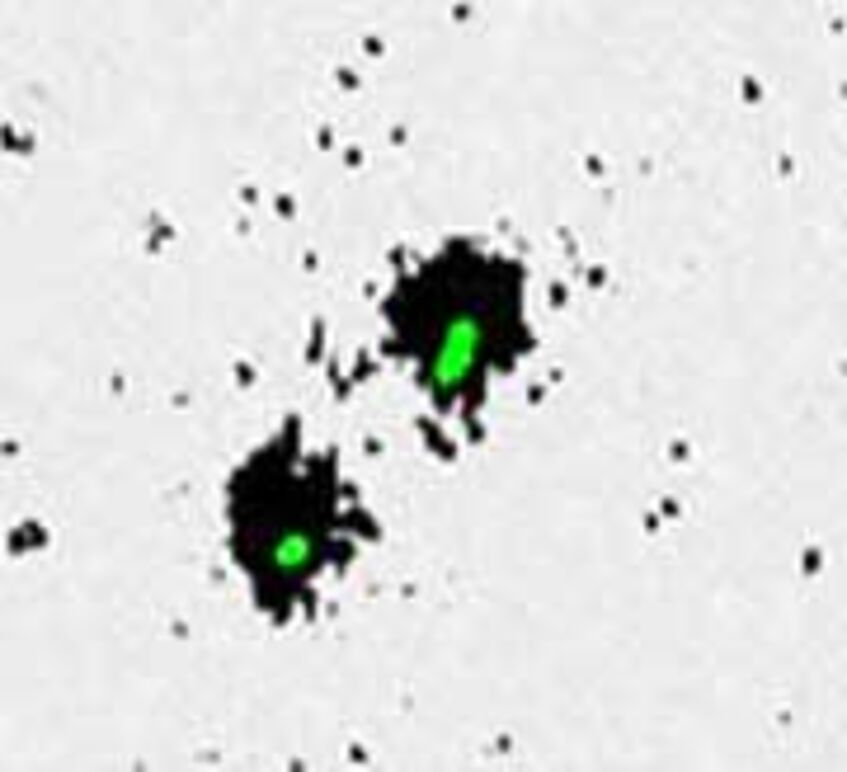
Due to rather large HRP-probes the permeabilization of the prokaryotic cell wall is crucial. While for bacteria lysozyme treatment is usually sufficient the archaeal cell wall has to be treated with proteinase K to allow penetration of the probes. With microautoradiography it is possible to visualize the uptake of radiolabeled substrates such as leucine or glucose by the prokaryotes. After the hybridization step the samples are developed similar to photographic film resulting in silver grain halos around those cells which incorporated the radioactive molecules. Thus in combination with CARD-FISH it is possible to get information on who are the main microbial groups in a water sample and which ones are predominantly active. We developed a thorough protocol specifically for open ocean prokaryotes which is available here:
Further information on the FISH technology can be found at on the SILVA database project webpage.
Suggested reading:
Teira, E., T. Reinthaler, A. Pernthaler, J. Pernthaler, and G. J. Herndl. 2004. Combining catalyzed reporter deposition-fluorescence in situ hybridization and microautoradiography to detect substrate utilization by bacteria and archaea in the deep ocean. Applied and Environmental Microbiology 70: 4411-4414.
Reprint (PDF)
Sintes, E., G.J. Herndl. 2006. Quantifying substrate uptake of individual cells of marine bacterioplankton by catalyzed reporter deposition fluorescence in situ hybridization combined with microautoradiography. Applied and Environmental Microbiology 72: 7022-7028.
Reprint (PDF)
Pernthaler, A., J. Pernthaler, and R. Amann. 2002. Fluorescence in situ hybridization and catalyzed reporter deposition for the identification of marine bacteria. Applied and Environmental Microbiology 68: 3094-3101.
Cottrell, M. T., and D. L. Kirchman. 2003. Contribution of major bacterial groups to bacterial biomass production (thymidine and leucine incorporation) in the Delaware estuary. Limnology and Oceanography 48: 168-178.
Ouverney, C. C., and J. A. Fuhrman. 1999. Combined microautoradiography 16S rRNA probe technique for determination of radioisotope uptake by specific microbial cell types in situ. Applied and Environmental Microbiology 65: 1746-1752.
Herndl, G.J., T. Reinthaler, E. Teira, H. van Aken, C. Veth, A. Pernthaler, J. Pernthaler, 2005. Contribution of Archaea to total prokaryotic production in the deep Atlantic Ocean. Applied and Environmental Microbiology 71: 2303-2309
Reprint (PDF)